





-

植物与我们的生活息息相关,虽然这个频道也做过有关植物的文章,但也只是草草带过,在这里我想专门开一个有关的频道,既为大家介绍日常生活中的息息相关的植物,也深入从科学的角度拓展冷知识,以不浮于表面看问题的态度来面对这个蓝色地球的真正原住民---植物。
我们先从最基础的科普知识说起,该篇相当于植物的简历,一份概括性文章,其他详细的,有趣的植物故事都是建立在该篇之上。
首先植物是植物界各式生物的统称,对于其范围,在历史上多有变动。在前林奈时代到林奈时代,除了现今所熟知的植物类群外,原核生物、真菌及藻类因不能主动运动或能进行光合作用等与植物类似的特性,也被划分进植物界内。而根据恩斯特·海克尔、罗伯特·魏泰克、琳·马古利斯、汤玛斯·卡弗利尔-史密斯等生物学家的进一步研究,原核生物、真菌和部分藻类先后被排除出植物界。在近年来的系统发育和分类学研究中,一个常被采用的定义是1981年汤玛斯·卡弗利尔-史密斯定义的植物界,将原始色素体生物视为与植物等同的概念;另一常见的观点则由赫伯特·科普兰在1956年定义、只包含绿色植物的植物界。

据估计,植物界现存大约有450000个物种,根据邱园的统计,直至2017年,在维管植物中,381910个物种已被描述和命名,其中369000种被子植物,1110种裸子植物,1300种石松类植物,10500种链束植物(狭义蕨类植物)。与此同时,也不断有新的物种被发现,例如在2016年,有1730种有胚植物被发现和命名。
有丝分裂和减数分裂是植物细胞主要的增殖方式,但无丝分裂在植物生长和发育中也有重要作用。有性生殖和世代交替是植物的重要特征,但无性繁殖也是重要的繁殖方式:。(以后会专门介绍几种增殖方式)
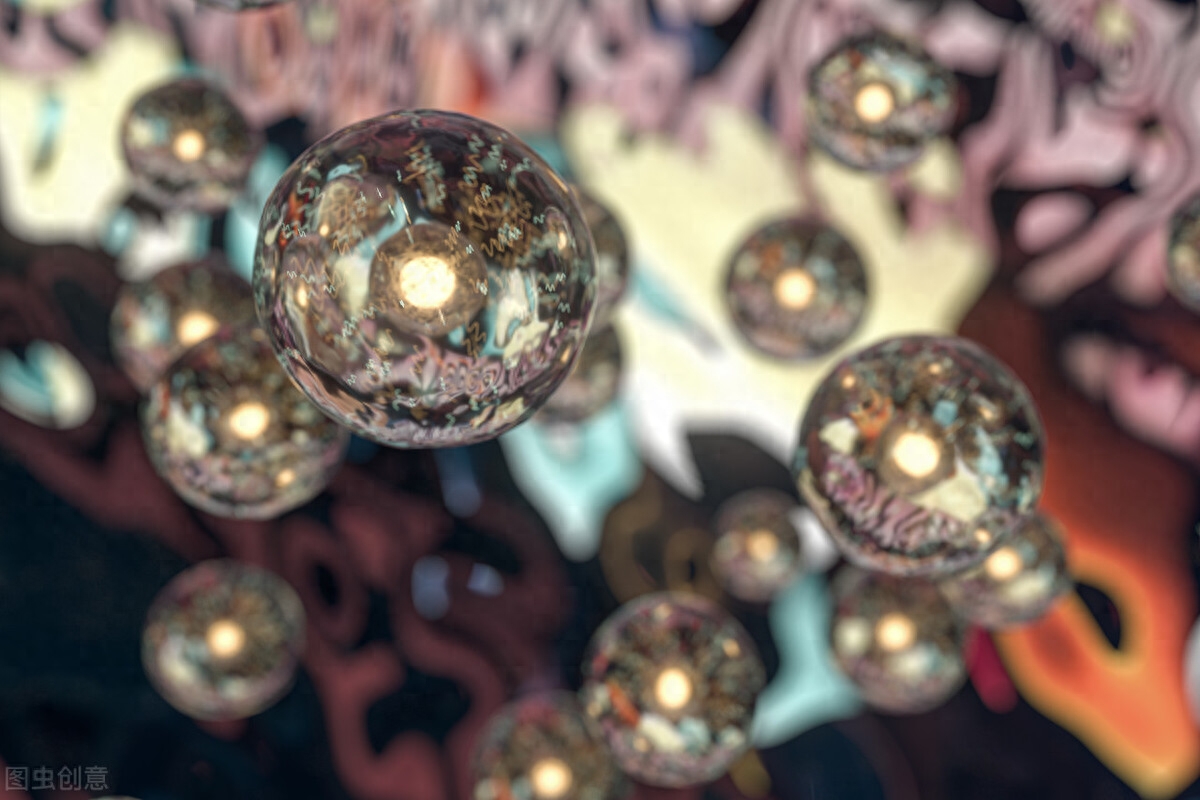
植物的共同祖先通过与蓝藻内共生获得了质体,并具备了光合作用的能力。绝大多数植物从祖先那里继承了这一结构及能力,但也有少数植物丧失了叶绿体这一结构和光合作用的能力,转为营寄生生活或腐生生活。
植物的光合作用是植物主要的物质和能量来源,也是生物圈物质循环和能量流动的重要环节,是地球大部分生态系统的基础。许多植物在人类的生产生活中作为食物、药物、建筑材料、纺织原料等,发挥着重要作用。以植物为研究,是生物学的重要分支。

定义
不论在学术意义上还是通俗意义上,“植物”的定义都众说纷纭,多有不同,以下简要叙述植物范围的历史变迁。
极广义植物界
极广义植物界指包含细菌、古菌、真菌,以及褐藻等与有胚植物远缘的藻类(不一定包含全部现代意义上的细菌、古菌、真菌和藻类,但只含蓝藻而不含其他细菌的不在此列)。
在林奈时代,系统的进化理论仍未产生,因此,在他两界系统中,采用了极广义的植物概念,将所有营固着生活、光合自养的生物定义为植物。恩格勒系统继承了林奈的植物界概念,将藻类、细菌、真菌都包括在内,《植物分科志要》在恩格勒生前发表10个版本,去世后由迪尔斯和梅尔希奥各主持一次修订,仍然维持了最初的植物界概念,将植物界分为17门。许多现代植物学课本仍将细菌、古菌、真菌和藻类包含在植物界的范围内进行讲解,并在植物界的分门上采用1964年第12版恩格勒系统,因此该系统仍有较大影响力。
排除原核生物的广义植物界
随着显微镜的发明和应用,生物学家发现了许多兼具传统动物和植物特点的生物,例如裸藻、甲藻等。因此,德国生物学家海克尔先后提出了,两个名称范围基本相同,都排除了原核生物、黏菌和部分藻类,并将其分入原生生物界。
1983年至2009年,在 两位科学家的先后主持下,第13版恩格勒系统完成修订,并于2012年开始出版。该系统在具体纲、目、科的设置方面吸收了大量分子生物学成果,但对于植物界整体范围的界定只排除了非蓝藻的细菌,仍包含了真菌和藻类。
排除真菌的较广义植物界
1959年,美国生物学家罗伯特·魏泰克将不含质体的广义真菌从植物界分出,提出了一个新的生物分类系统,并在1969年再次修订,称为“魏泰克五界系统”,该系统所定义的植物界;排除了原核生物、真菌和部分藻类,将其分别分入原核生物界、真菌界和原生生物界,但仍包含与陆生植物关系较远的褐藻类。

内共生理论与原始色素体生物
1966年,琳 · 马古利斯发表论文,阐述了著名的内共生理论,该理论认为,真核生物的共同祖先通过内共生某种好氧细菌获得了线粒体和有氧呼吸的能力,而后植物的共同祖先通过内共生某种蓝藻而获得了含叶绿体在内的质体和光合作用的能力。
马古利斯在同一篇论文中定义其为维管植物,而后,马古利斯又在1971年将她定义的植物界 的范围改为有胚植物。
内共生理论对于分类学的最重大影响在于,在该理论下,核基因组和细胞器基因组的演化历史并不一致,因此可以通过二者的差别来判断内共生的发生。英国牛津大学汤玛斯·卡弗利尔-史密斯研究组的分子系统学的结果显示,原始色素体生物(包括灰藻、红藻、绿藻、轮藻和有胚植物)是最早与蓝藻内共生获得质体的生物类群,其他藻类的质体都并非直接来自蓝藻,而是通过与原始色素体生物内共生而间接地获得质体,该研究组还定义为原始色素体生物。
在近年来的系统发育和分类学研究中,普遍采用将原始色素体生物视为与植物等同的概念。
新的争议
2015年以来,汤玛斯·卡弗利尔-史密斯、帕特里克 J. 基林各自所带领的研究组的研究对现有的植物定义提出了挑战。
前三者的分子系统学的结果均显示,一类原生生物皮胆虫嵌在了原始色素体生物的演化支中,并与红藻类构成姊妹群。

隐藻生物的分类地位尚不明确,帕特里克 J. 基林研究组的系统树均显示,隐藻生物可能嵌在原始色素体生物中,但其具体位置则并不一致;在前者的结果中,灰藻和绿色植物构成单系,隐藻则是它们的旁系群;在后者的结果中,隐藻与红藻互为旁系群;而另两个研究组的系统树则显示,隐藻生物可能是原始色素体生物和皮胆虫的外类群。
这些研究显示,皮胆虫和隐藻很可能是原始色素体生物共同祖先的后代,但其各自在演化过程中将祖先的质体丢失,而后隐藻通过内共生获得了红藻的质体,但皮胆虫则恢复为异养生活。
多样性
植物是地球上最常见的生命形态之一,广泛分布于各类生境中,包括全世界水圈的大部,岩石圈的表面,大气层的底部,随着不同气候区而有不同的种类和数量。甚至在沙漠、高原、极地等极端环境下也有植物的身影。
植物中的绿藻可与真菌共生形成地衣,并共同在一些极端环境下生存。地球陆地面积的6%被地衣覆盖,北极地区的地衣是北极驯鹿的主要食物,某些地衣甚至可以抵挡高强度的宇宙射线与紫外线,在人造卫星上的研究站存活。
据估计,植物界现存大约有450000个物种,根据邱园的统计,直至2017年,在维管植物中,381910个物种已被描述和命名,其中369000种被子植物,1110种裸子植物,1300种石松类植物,10500种链束植物(狭义蕨类植物)。其他类群在邱园的数据中未被统计,其物种数目及参考资料详见下表。
与此同时,也不断有新的物种被发现,例如在2016年,有1730种新发现的有胚植物被命名。
演化史
依照目前较广泛接受的研究,植物(原始色素体生物)起源于16亿年前的一次内共生事件,其中的质体、线粒体等结构,则可以追溯到更早存在的蓝藻和好氧细菌。
蓝藻的历史没有定论,最早的可能记录是超过35亿年前的蓝藻黏结堆砌而成的叠层石,但仍有争议。在最晚16亿年前,植物的最后共同祖先以蓝藻为食,但在食物缺乏且阳光充足的环境下,与蓝藻共生,利用它们进行光合作用比将蓝藻作为食物更有利于生存,因此植物的祖先与蓝藻发生了内共生,并获得了质体和光合作用的能力,而一种类似衣原体的生物(可能是军团菌,包括引发军团病的种类)则提供了将有机物转运出质体的基因。
传统上认为有胚植物是最早的陆生植物,但近年来有研究指出,通过对古土壤的化学和地质学分析可以看出,早在10亿年前就有陆生单细胞藻类。
有胚植物起源于轮藻,最早产生于奥陶纪中期,传统观点认为其起源于海洋藻类,但也有研究指出其起源于陆生单细胞藻类;维管植物出现于志留纪时,且到了泥盆纪时已辐射并分布在许多不同类型的陆地生态系统中;早期种子植物——种子蕨和科达树出现于泥盆纪晚期并于石炭纪时开始分歧,之后经过二叠纪和三叠纪而有了更多的演化;被子植物是最后出现的植物类群,于侏罗纪时由古裸子植物中演化出来,并于白垩纪时迅速地分歧出许多物种,并一直繁盛至今,成为陆生植物的最优势类群;现代裸子植物(称为末端裸子植物)是与被子植物并列的独立单系分支,并非被子植物的祖先。

化石记录
主条目:古植物学
植物的化石包括根、茎、叶、种子、果实、花粉、孢子和琥珀。植物化石在陆地上、湖泊中、河流里以及近海内的地层都有发现的记录。花粉、孢子和藻类的化石被用来界定地层岩石的顺序。残留的植物化石并不如动物化石那么普遍,然而植物化石在世界上许多地区之内,都可以有大量的发现。

石化林国家公园中的硅化木。
最早可以被明确归类于植物的化石是一种红藻,生活在16亿年前,发现于印度的叠层石中。
绿色植物最早的化石是澳大利亚卢坡群(Roper Group)的一些微体化石,生活在15亿年-13亿年前,其结构与现代绿藻类似。
最早的与现生类群有明确亲缘关系的植物化石是一种生活在12亿年前的红藻。
轮藻门的一些植物的细胞壁发生钙化,因此比其他藻类更容易留下化石记录,可追溯到志留纪晚期(大约4.1亿年前),例如派卡藻等。

有胚植物已知最古老的遗迹化石源于奥陶纪,但此类化石是零碎不全的。到了志留纪才有完整的化石被保留下来,包括石松门的“刺石松”。泥盆纪之后,莱尼蕨门的详细化石在此时期被发现,此化石显示出了其植物组织的单一细胞。泥盆纪时亦出现了被认为是最古老树木的植物“古羊齿属”,此类植物在其树干上有蕨叶,但此蕨叶不会产生孢子。
煤系地层是生存于古生代时的植物的化石的主要来源。煤炭矿是采集化石最好的场所,而且碳本身便是化石植物的残留,虽然植物化石的结构细节很少会留在碳中。在苏格兰格拉斯哥维多利亚公园中的化石森林里,有发现鳞木(Lepidodendron)的树干。
松柏和开花植物的根、茎及枝干的化石可以在湖泊及海岸的中生代和新生代地层中被找到。加州红木、木兰、栎树和棕榈树等化石常被找到。
石化木普遍存在于世界的部分地区,且最常在酸性及沙漠地区中所发现,因为那些地方很容易因侵蚀作用而暴露。石化木通常都被严重地硅化(有机质被二氧化硅取代),且生殖组织常会被保存在良好的状态。此类的样本有些会使用宝石雕琢的装置来切割及磨光。石化木的石化林已在每个大陆中被发现。

舌羊齿属等种子蕨的化石广泛分布在南半球的数个大陆中,此一事实支持了阿尔弗雷德·魏格纳所提出的大陆漂移学说。
目前无争议的最早的化石被子植物是古果属植物;最早的与现生类群亲近的化石被子植物是蒙特塞克藻,其与金鱼藻科互为姊妹类群。
另有潘氏真花等位置未定的古种子植物,部分学者主张其为被子植物,也有学者质疑其为松柏类裸子植物。若干项关于被子植物起源时间的研究认为,依据分子钟的结论,被子植物可能起源于侏罗纪甚至三叠纪,但尚无可靠的化石证据支持,分析指出,这可能是由于早期被子植物在生态上并不重要,或者其生活在化石化作用难以发生的环境中。

外类群
依据目前的研究,植物界是双鞭毛生物>多毛生物的一支,其与相关类群间的系统发生关系如下
分类
植物分类学史
主条目:植物分类学
植物分类学是一门历史悠久的学科,古希腊哲学家、科学家亚里士多德将植物分为乔木、灌木和草本三大类,他的学生、植物学鼻祖泰奥弗拉斯托斯在《植物的历史》和《植物本原》两部著作中记载了500多种植物,在这些著作中,他按照生殖方式、生境、植株大小、食用、药用价值等对植物进行分类。在古代中国,嵇含的《南方草木状》、李时珍的《本草纲目》、吴其窃的《植物名实图考》等著作,也是根据生境、生活型、生态型、生活生产用途等特征进行分类。由于这些著作中的分类系统反映的都是人们对植物的认识和使用,而非不同植物之间的亲缘关系,因此它们都是典型的人为分类系统。
人为分类学的巅峰人物是现代生物分类体系的鼻祖卡尔·冯·林奈,他在《植物种志》中,在前人成果的基础上,提出了以双名法及阶元体系为核心的林奈氏分类法,并为他当时所知的物种提出了双名法形式的学名。林奈提出了“性系统”,依据花器官的数量等特征为植物分门别类,将植物分为了单雄蕊纲、双雄蕊纲等。
1858年,查尔斯·达尔文和阿尔弗雷德·拉塞尔·华莱士提出了生物的进化论,在此影响下,新的观念被引入了分类学界,也即,生物的分类系统应当反映出类群间的亲缘关系和系统发生过程,这一阶段分类学的主流思想是进化分类学,相比于林奈时代“上帝创造,林奈整理”的神创论和物种不变论的思想,有了较大的进步。相对于古代和林奈时代的人为的分类学,我们将进化分类学称为“自然”的分类学,但是这一阶段主要的证据来自于表型,证据少、主观性强、依赖于分类学家个人的经验和权威、并难以排除趋同演化的影响;而且进化分类学派承认并系群的合理性,而并系群并不是真正的自然类群,这些都使得进化分类学并不是真正自然的分类学。
德国昆虫学家维利·亨尼希于1965年提出了亲缘分支分类学,也称支序分类学,他主张,只有单系群才是自然类群,才是合理的分类群,任何一个分类群都应该包含其最后共同祖先的全部后代,在实际操作上,应当根据祖征和衍征,按照简约化的方式确定亲缘关系,并依此进行分类。
随着分子生物学的产生与发展和测序技术的成熟,分子生物学证据逐渐取代了形态学证据成为了系统树主要的数据来源,由于分子生物学证据信息量大,主要的构建算法(邻接矩阵法、贝叶斯法、最大简约法、最大似然法等)计算量庞大,人力难以准确完成,因此现今一般使用计算机软件进行计算。
藻类
、

主条目:藻类
藻类传统上被视为植物的一部分,但分子生物学研究显示其不是一个自然类群。藻类是由若干种可以进行光合作用的不同类群生物所组成的,但大多数的藻类并不被归类在植物界里,而是被归类在细菌界(蓝藻)、SAR超类群(金藻、褐藻、黄藻、硅藻、甲藻)、定鞭藻门和古虫界(裸藻)里。
有胚植物是由轮藻演化而来的;二者被合称为链形植物,加上绿藻门共同组成绿色植物。传统上绿藻不包含有胚植物,因此为一并系群,但后来因为许多研究显示有胚植物是由轮藻门的一支演化而来,部分研究者开始将两者视为一体。这些植物的细胞壁中存在纤维素,质体由内共生的蓝藻演化而来,含有叶绿素 a 以及叶绿素 b,不含藻胆素,该类群以没有中心粒的方式行有丝分裂,且具有线粒体。
红藻具有叶绿素 a 和藻胆蛋白,类似于大多数蓝藻。灰藻具有典型的蓝藻色素,并且其色素体保留了蓝藻的肽聚糖壁,称为蓝小体 。
绿藻、红藻和灰藻是最早与蓝藻内共生获得质体的生物类群,它们(包括由绿藻演化来的整个绿色植物)因此被称作“原始色素体生物”。相对的,大多数其他藻类的质体则被三至四层细胞器膜包裹,依据核基因组来看,它们和植物的关系并不相近,可能是将绿藻和红藻内共生至自己的细胞内而得来的。
后文提及“藻类”,如非特别说明,均指原始色素体生物中排除陆生有胚植物的部分。
有胚植物[编辑]
主条目:有胚植物
软树蕨,树蕨的一种物种。

生活中常见的多细胞陆生植物为有胚植物。根据是否拥有完整的维管组织,可以划分为拥有维管组织的维管植物(但松叶蕨科缺少真正的根)以及缺少维管组织的苔藓植物。
这类植物都有着以纤维素、半纤维素、果胶等成分组成的细胞壁所包围的真核细胞,且大部分经由光合作用来合成营养物质。也有许多植物不行光合作用,而是寄生在其他生物体上,在被子植物中,寄生植物约有4100种。有胚植物和藻类不同,有着被非生殖组织保护着的特化生殖器官。
苔藓植物中,尽管有些物种相对耐旱,但其繁殖离不开水,大多数的苔藓植物一生的体形都很小,它们在两个阶段之间做世代交替:称为配子体的单倍体阶段和称为孢子体的双倍体阶段。孢子体的生存期间很短,且必须生活在雌配子体上。
维管植物有很多种适应的型态,允许它们能够克服苔藓植物的极限。角质层能帮植物对抗干燥,而维管束则能把水分输送到整个生物体内。维管植物的孢子体一般是独立的植物个体,而配子体只作为生殖结构短期出现。
种子植物的配子体阶段完全地退化了,而其孢子体则是通过花粉受精,在母本体内发育成种子。种子植物可以在极端干旱的条件下存活和繁殖,因为它们能够形成花粉管,受精过程不依赖水,也不依赖于配子体的独立生活。
早期的种子植物为裸子植物,其胚珠不被大孢子叶所形成的心皮包被,种子没有果皮包被。现代裸子植物依然广泛分布于世界各处。

而被子植物的孢子体进一步进化,配子体进一步退化,胚珠被心皮形成的雌蕊包被,种子被果皮包被,形成果实,具有真正的花,并能通过双受精形成具有双亲遗传物质的胚乳。在现今大多数的生态环境里,被子植物是最主要的类群。
形态结构
主条目:植物形态学
植物形态学是一门研究植物的外部形态、构造、组成,及其发育(包括个体发育与系统发育)的科学。
细胞
植物作为真核生物,其细胞具有真核细胞的共同特征:被核膜包裹的真正的细胞核和具膜细胞器。
植物细胞由细胞壁和原生质体组成,原生质体又可分为细胞膜、细胞质和细胞核。
细胞膜又称原生质膜,由磷脂双分子层和镶嵌在其上的糖蛋白构成,它将原生质体与外部环境分离,并能介导物质进出原生质体。
细胞质包括细胞质基质和细胞器,植物细胞中有无膜结构的细胞器如核糖体等,也有双层膜细胞器如质体、线粒体,单层膜细胞器如内质网、高尔基体、溶酶体、液泡;核膜、内质网、高尔基体、溶酶体、液泡等的膜结构共同组成植物细胞的内膜系统。在活的植物细胞中,细胞质总是在运动,可以观察到细胞器以及悬浮在细胞质基质中的各种物质在有序地运动,这种运动被称为细胞质环流。
细胞核由核膜、核仁、核基质、染色质、核纤层、核膜核糖体等部分组成,它通过控制细胞何时产生何种蛋白质来控制细胞的生命活动;它将遗传信息以DNA储存在其染色质中,在细胞分裂时其传递给子细胞。存储在细胞核中的全部遗传信息称为核基因组。

质体(质体基因组)和线粒体(线粒体基因组)中也含有DNA,储存了部分遗传信息。
植物细胞的细胞壁由纤维素、半纤维素、果胶质等化学物质组成,细胞壁可分为三层,胞间层是细胞壁的最外层,是相邻细胞共有的一层结构,在分裂过程中最早形成;初生壁在胞间层的内侧,初生壁和胞间层都较为柔软,可随着细胞生长而扩展;次生壁在细胞体积停止增大后形成,含有木质素,一般在起支持和运输作用的细胞中见到,并非所有细胞都有次生壁。
细胞的增殖
细胞的增殖方式称为细胞分裂。作为真核生物,植物的细胞增殖方式与大部分真核生物一样,以有丝分裂为主要的体细胞分裂方式,兼有无丝分裂;以减数分裂作为产生生殖细胞的分裂方式。植物的有丝分裂和动物一样,是开放式有丝分裂,而真菌和一些原生生物(包括一些非植物藻类)则进行封闭式有丝分裂。
参见:细胞分裂、有丝分裂、无丝分裂和减数分裂
组织
植物的组织主要包括基本组织、维管组织和皮组织(也称为保护组织)。
基本组织又可分为薄壁组织、厚角组织和厚壁组织,后两者也常被合称为机械组织。薄壁组织是最常见的基本组织,只有初生壁,在光合作用、储水、传递等方面发挥作用;薄壁组织具有全能性,它们与伤口的愈合、不定根的发生也有着密切的关系。
厚角组织和厚壁组织起支持作用,前者的细胞壁部分加厚,常出现在茎、叶柄等部位,是活细胞;后者的细胞壁全面加厚,具有次生壁,是死细胞,由两类细胞组成:纤维和石细胞。
维管组织包括木质部和韧皮部,木质部包括导管、管胞(少数被子植物、裸子植物和蕨类植物)、木纤维和木薄壁细胞,韧皮部包括筛管、筛胞(裸子植物和蕨类植物)、伴胞、韧皮纤维和韧皮薄壁细胞;植物通过木质部将根部吸收到的水分和矿物质向上运输,通过韧皮部将光合作用的产物向下运输。
皮组织也称为保护组织,包括表皮和周皮。表皮在最外层,是初生保护组织,存在于叶、花、果实、种子以及未成熟的茎和根的表面。周皮是次生保护组织,存在于成熟的茎和根中,代替表皮起保护作用。
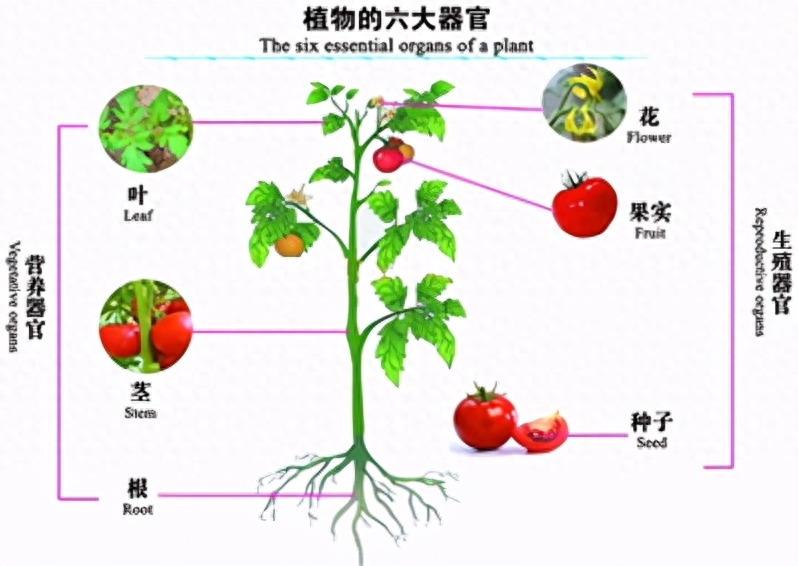
器官
植物的器官包括根、茎、叶三大营养器官和花、果实、种子三大生殖器官,其形态和结构是传统植物形态学的研究重点,相关特征常作为传统植物分类学上区分物种的证据,尽管分子生物学证据在现代有着更广泛的应用,但在野外等不方便采用分子生物学手段的场合,形态学证据仍然有着广泛的应用。
植物的根通常位于土壤中,主要功能为吸收、输导和固着支持,气生根、贮藏根等变态根还具有合成、贮藏、呼吸等功能。真正的根最早在维管植物中产生,苔藓植物和藻类也有类似的结构,称为假根,只起固着作用,不具备其他功能。种子植物的主根由种子中的胚根发育形成,主根垂直向下扎入土壤中,长到一定长度后,从主根的中柱鞘结构产生侧根,侧根上还可以进一步生出更多侧根,在主根和侧根以外的部分如茎、叶、胚轴或老根上产生的根称为不定根。
植物的茎通常位于地面以上,向下与根相连,向上着生叶、花、果实,具有输导、支持的功能。茎具有节和节间,节是叶和芽着生的位置,芽可以根据位置分为顶芽和侧芽,也可以根据将来发育成的器官分为枝芽、花芽和混合芽。
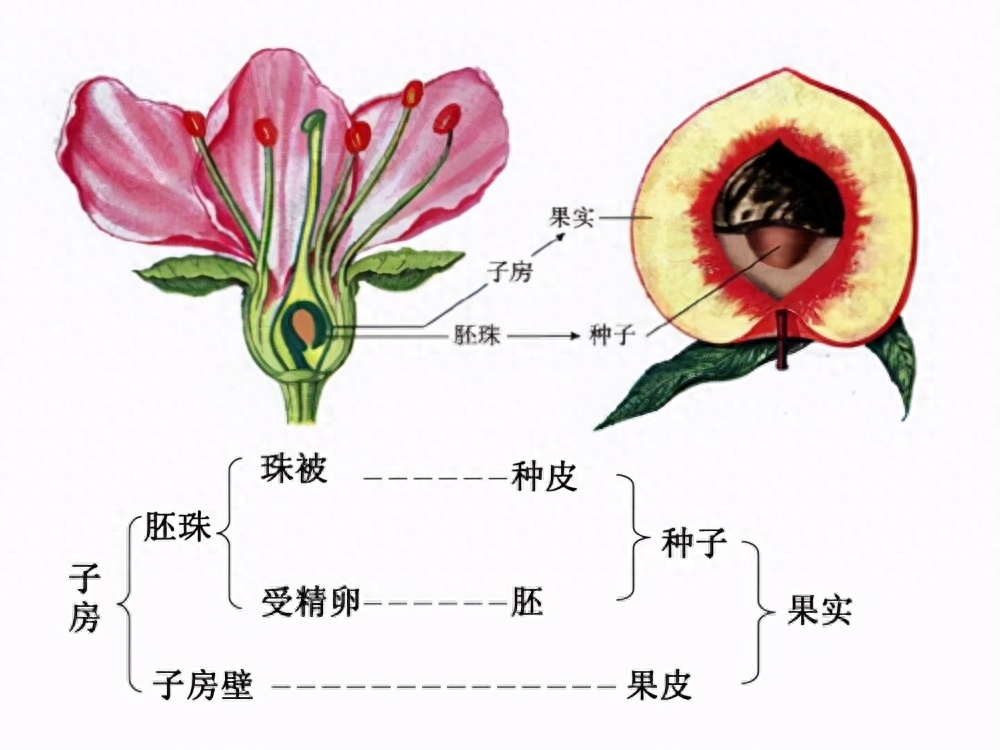
叶是植物行使光合作用的主要部位。
植物的叶是光合作用制造有机物的营养器官,其主要功能是光合作用和蒸腾作用。大部分真双子叶植物和木兰类植物的叶可以分为叶片、叶柄和托叶三部分;大部分单子叶植物和一部分真双子叶植物,叶基部扩大,形成包裹茎的叶鞘;在禾本科植物中,叶鞘会延伸节间的长度。对大部分植物而言,叶片是最主要的部分,一般为绿色薄片状,有利于扩大阳光照射面积和气体接触面积,以便于光合作用和蒸腾作用的进行。
变态的茎和叶还具有其他功能,例如马铃薯的块茎,洋葱的鳞茎,唐菖蒲的球茎均具有贮藏功能,龙舌兰属植物的茎、景天属植物的叶则具有储水功能。
真正的茎和叶最早产生于石松植物中,而类似茎和叶的结构早在苔藓植物中已经出现,但因为其不具有维管组织,因此不能算做真正的茎和叶。其中只有叶状结构的,称为“叶状体”,具有茎状和叶状结构的,称为“拟茎叶体”。
植物的根、茎、叶都具有韧皮部和木质部,并在各个器官中相互连通。
种子只存在于种子植物中,而花、果实则只在被子植物中存在。苔藓、蕨类植物一般使用孢子繁殖;而藻类除孢子繁殖外,也进行营养生殖、接合生殖等。
繁殖与生活史
植物界是一个古老而庞大的类群,演化出了许多不同的繁殖方式和生活史,大部分多细胞植物的生活史具有世代交替现象,但这一现象在不同类群中也不相同,少数不具备世代交替,仅具备核相交替。以下依类群简要介绍。
灰藻
灰藻行开放式有丝分裂,通过产生游动孢子或非运动的似亲孢子来进行无性繁殖。尚未发现灰藻能够进行有性生殖的证据。
红藻
红藻的生殖方式有无性生殖、有性生殖。
在无性生殖中,红藻将孢子排放到水中,如果孢子遇到合适的基质,就将附着在其上开始生长。
红藻的有性生殖生活史较为复杂。较简单的一种生活史涉及到单倍的配子体和二倍的孢子体两个阶段,配子体上,雄性生殖结构产生的雄配子是不动精子,不具备鞭毛,借助水流移动;红藻的雌性生殖结构称为果孢,其上具有称为受精丝的突起,用于接受精子。精子接触到受精丝并结合后,雄核经由受精丝向下移动到雌核并与之融合,形成的二倍体受精卵,受精卵有丝分裂产生果孢子。果孢子从亲本配子体释放到水中,附着在某处生长为孢子体,孢子体通过孢子减数分裂产生单倍体孢子。单倍体孢子又会发育为配子体,从而完成孢子体-配子体两个世代的世代交替。
大部分红藻的生活史具有三个世代的交替,这种生活史与上一种生活史的主要区别在于,精子和卵细胞结合后,受精卵并不直接分裂产生果孢子,而是通过反复进行有丝分裂,产生第三个世代,二倍体的果孢子体。果孢子体并不独立生活,只附着在亲本配子体上,并从配子体上吸收养分进行生长,果孢子体成熟后,进行有丝分裂产生果孢子,并从果孢子体释放到水中,附着在某处生长为二倍体的四分孢子体,再进行减数分裂,产生四分孢子,发育成配子体。这些红藻从而完成了配子体-果孢子体-四分孢子体三个世代的世代交替。
红藻的孢子体或四分孢子体与配子体的形态差别不大,而果孢子体与二者差别较大。
绿藻及轮藻
绿藻及轮藻的繁殖方式多样,有无性生殖、有性生殖,又因单细胞植物体、多细胞植物体、生殖结构是否被营养结构包被而有不同;还具有接合生殖等特有的生殖方式。
一种轮藻的生殖结构,上方的结构为藏卵器,下方的结构为藏精器。
一些单细胞绿藻既能进行无性生殖,也能进行有性生殖,衣藻属是其典型代表。无性生殖时,单倍体核行有丝分裂,在细胞壁内产生最多16个子细胞。然后每个子细胞各自的细胞壁和鞭毛,并分泌一种分解亲代细胞壁的酶,这些子细胞——游动孢子释放出来,各自发育成新个体。有性生殖时,衣藻的营养细胞在缺氮条件下形成配子,配子相互结合,形成二倍体的受精卵。受精卵周围形成厚厚的细胞壁,经过一段时间的休眠后,受精卵进行减数分裂,产生四个单倍体细胞,这些细胞可以进行分裂或相互交配以产生新的受精卵。
石莼属是多细胞植物,藻体叶片状,石莼属的营养结构和生殖结构没有绝对的界限,除基部及固着器以外,孢子体每个细胞均可形成孢子囊,其内的细胞核进行减数分裂,产生多个单倍体的具有4条鞭毛的游动孢子。游动孢子释放出来后,可以萌发产生单倍体的配子体。除基部及固着器以外,配子体成熟时,每个细胞均可形成配子囊,经有丝分裂产生多个游动配子(+配子和-配子同型),配子结合后形成合子,合子萌发产生孢子体。石莼属的孢子体和配子体形态上区别不大。
轮藻门中一些相对复杂的种类具有较为复杂的生殖结构:产生精子的藏精器和产生卵细胞的藏卵器,这两种结构生在节上,对于雌雄同体的轮藻而言,藏卵器在上方,藏精器在下方。轮藻的生殖结构中,生殖细胞被外侧的营养细胞包被保护,藏卵器的外侧有若干长而呈管状、扭曲的细胞保护内部的卵细胞,而藏精器的外部则由若干盾形细胞保护。精子是轮藻生命周期中唯一具有鞭毛的细胞,其形成后,穿过藏卵器的保护结构与卵子结合成受精卵,合子经过休眠后进行减数分裂产生四个子细胞,其中仅有一个发育为新个体。
苔藓植物
蕨类植物
裸子植物
被子植物
生态
主条目:植物生态学
陆生植物和藻类所行使的光合作用几乎是所有的生态系中能源及有机物质的最初来源。光合作用根本地改变了早期地球大气的组成,使得现在有21%的氧气。动物和大多数其他生物是好氧的,依靠氧气生存。植物在大多数的陆地生态系中属于生产者,形成食物链的基本。许多动物依靠着植物做为其居所、以及氧气和食物的提供者。
植物的光合作用是生物圈物质循环和能量流动的重要环节,是地球大部分生态系统的基础,一些植物(如豆科植物等)和固氮菌共生,使得植物成为氮循环重要的部分。植物根部在土壤发育和防止水土流失上也扮演着很重要的角色。植物通常是它们栖所上主要的物理及结构组成。许多类型的形态系统即以植被的类型而命名,如草原和森林等等。它们通过遗传分化和表型可塑性来适应不同环境。
根据邱园的统计,植物和鸟类的生物多样性区域有53%的重叠。
生态关系
捕蝇草-一种食虫植物
许多动物和植物共演化,例如:许多动物会帮助花授粉以交换其花蜜;许多动物会在吃掉果实且排泄出种子时帮到植物散播其种子。适蚁植物是一种和蚂蚁共演化的植物。此类植物会提供蚂蚁居所,有时还有食物。做为交换,蚂蚁则会帮助植物防卫[[帮助其和其他植物竞争。蚂蚁的废物还可以提供给植物做有机肥料。
大部分植物的根系会和不同的真菌有互利共生的关系,称之为菌根。真菌会帮助植物从土壤中获得水分和矿物质,而植物则会提供真菌从光合作用中组成的碳水化合物。一些植物会提供内生真菌居所,而真菌则会产生毒素以保护植物不被草食性动物食用。[15]:684植物的根系也可与细菌共生,形成根瘤,是根上的瘤状突起,如豆科植物、木麻黄、罗汉松、杨梅等,能够与土壤中的根瘤菌共生,根瘤菌能将空气中的氮转变为氨,提供给植物,同时根瘤菌可以从植物的根中吸取水分和养料,供自身生长发育所需。[15]:693
许多种类型的寄生在植物中亦是很普遍的,从半寄生的槲寄生(只是从其寄主中得取一些养分,但依然留有光合作用的叶子)到全寄生的列当和齿鳞草(全部都经由和其他植物根部的连结来获取养分,所以没有叶绿素)。一些植物会寄生在菌根真菌上,称之为菌根异养,且因此会像是外寄生在其他植物上。
许多植物是附生植物,即长在其他植物(通常是树木)上,而没有寄生在其上头。附生植物可能被间接地伤害到其宿者,经由截取宿者本应得的矿物质和太阳光。大量附生植物的重量可能被折断树干。许多兰花、凤梨、蕨类植物和苔藓通常会是附生植物。凤梨科的附生植物会在其叶腋上累积水分而形成树上水池,一种复杂的水生食物链。
少部分植物是食虫植物,如捕蝇草和毛毡苔。它们捕捉及消化小动物以获取矿物质,尤其是氮。

保护
由于人类的大规模活动,造成了许多全球性的环境问题,例如温室效应、全球变暖等,使许多植物面临绝灭的危险。国际植物遗传资源委员会(IBPGB)为此建立了国际基因库联网中心,贮存更多的植物基因。
与人类的关系
研究植物对人类的用途的学科被称之为经济植物学或民族植物学。这两个词通常被当做同义词,但有些人认为经济植物学主要专注于对现今作用的用途,而民族植物学主要则是在研究当地住民对其本土植物的应用。人类对植物的栽种是农业的一部分,其为人类文明的基础。植物农业可分成农学、园艺学和林业。
食物
实际上,所有人类的养分来源都直接或间接地依靠着陆生植物。绝大多数的人类的养分依靠谷物,尤其是玉米、小麦和稻米,或者是其他主食如马铃薯、木薯和荚果等。其他被食用的植物部分还包括水果、蔬菜、坚果、香草、香料和食用花卉等。由植物制成的饮料包括咖啡、茶、葡萄酒、啤酒等。糖主要是由甘蔗和甜菜中得到的。食用油和植物牛油来自玉米、大豆、芥花籽油、红花、向日葵、橄榄等等。食品添加剂包括阿拉伯树胶、瓜尔胶、刺槐豆胶、淀粉和果胶等。
非食用性产品

毛地黄
木材被用在建筑、家具、纸张、乐器和运动用具上头。布料通常是由棉花、亚麻或其原料为纤维素的合成纤维,如嫘萦和醋酸根。来自植物的可再生燃料包括柴、泥炭和其他生质燃料。炭和石油是来自于植物的化石燃料。来自于植物的药物包括阿司匹灵、紫杉醇、吗啡、奎宁、利血平、秋水仙素、毛地黄和长春新碱等。植物中存在于上百种药草如银杏、紫锥花、解热菊和贯叶连翘等。来自于植物的农药包括尼古丁、鱼藤酮、番木鳖碱和除虫菊精类等。来自于植物的毒品包括鸦片、古柯碱和大麻等。来自于植物的毒药包括蓖麻毒素、毒参和箭毒等。植物是许多天然产品如纤维、香精油、染料、颜料、蜡、丹宁、乳胶、树脂、松香、生物碱、琥珀和软木的源料。源自于植物的产品包括肥皂、油漆、洗发精、香油、化妆品、松节油、橡胶、亮光漆、润滑油、亚麻油地毡、塑胶、墨水、口香糖和麻绳等。植物亦为大量有机化合物的工业合成中,基本化合物的主要来源。
美观用途
成千的植物物种被种植用来美化环境、提供绿荫、调整温度、降低风速、减少噪音、提供隐私和防止水土流失。人们会在室内放置切花、干燥花和室内盆栽,室外则会设置草坪、荫树、观景树、灌木、藤蔓、多年生草本植物和花坛花草植物的意像通常被使用于美术、建筑、性情、语言、照像、纺织、钱币、邮票、旗帜和臂章上头。活植物的艺术类型包括绿雕、盆景、插花和树墙等。观赏植物有时会影响到历史,如郁金香狂热。植物是每年有数十亿美元的旅游产业的基本,包括到植物园、历史园林、国家公园、郁金香花田、雨林以及有多彩秋叶的森林等地的旅行。

文化
植物也为人类的精神生活提供基础需要。每天使用的纸就是用植物制作的。一些具有芬芳物质的植物则被人类制作成香水、香精等各种化妆品。
许多乐器也是由植物制作而成。而花卉等植物更是成为装点人类生活空间的观赏植物。

恭喜你。能看到这里说明你不仅是喜欢植物还是一个有耐心的人,也很高兴你能看到我的文章,如果觉得还不错的,别忘了点个赞,转发给你志同道合的朋友哦!
后续【科学馆】推出的植物文章将会更精细化,当然我会尽我所能的将其讲得通俗易懂。点关注,不迷路哦!
文章标签: 顶: 0踩: 0本文链接:https://www.lezhuanwang.net/kepu/77415.html『转载请注明出处』
相关文章
- 2023-09-02狮子头的家常做法(红烧狮子头的正确做法)
- 2023-09-02洛阳和西安哪一个才是第一古都(第一古都城市简介)
- 2023-09-02568b和568a有什么区别(网线水晶头的568A和568B标准异同)
- 2023-09-02荣耀x30和x40哪个好(荣耀X30和荣耀X40区别)
- 2023-09-01乌镇东栅和西栅有什么区别(乌镇西栅和东栅的不同之处)
- 2023-09-01武汉和武昌有什么区别()汉口汉阳武昌和武汉的关系)